As part of my ongoing travels around Glasgow’s green spaces, last weekend I headed southwest to Linn Park. I took the slightly longer, more scenic route, following the White Cart Water through a strip of deciduous woodland.
Whilst wandering along the shaded footpath, I noticed several small invertebrates on the wooden railings. They were little (around 3 mm long), dark, and surprisingly shiny. It wasn’t until I looked through my camera that I realised they were globular springtails – but not a species I recognised. I was also struck by how docile they were. Whenever I’ve tried to photograph springtails before, they’ve usually disappeared in a split second, but these seemed quite content to let me take a few photos without making much effort to escape.
After doing some online investigation that evening, I concluded that they were likely Allacma fusca. Described as large (for a globular springtail), dark brown, and with a 4th antennal segment which is divided into 16 subsegments. That did seem to check out, but as I hadn’t previously encountered this species, I remained unsure. I added my tentative ID to iRecord and the following morning checked the Glasgow Museums Biological Records Centre’s database. To my surprise, there were no records of this species in the GMBRC recording area, and when I checked NBN Atlas Scotland, I only found a limited number of records for the whole country.
More determined now to discover who these lovely little creatures were, I emailed James McCulloch, scheme organiser for the Springtail Recording Scheme. I got a prompt reply confirming that these were indeed Allacma fusca, and a suggestion that this species may actually be more abundant in deciduous woods in the Scottish lowlands than is currently known.
So it would be worthwhile, whenever you find yourself in this kind of habitat (especially in damper areas), to take a look and see if you can find this springtail. There’s a very good chance that it could be the first record for the site, or even the county.
Last week (22 – 28 June 2026), both the Royal Entomological Society‘s annual Insect Week and the GB Non-native Species Secretariat (NNSS) & Defra’s Invasive Species Week happened to coincide. As such, there was a series of excellent events and activities across Britain which aimed to celebrate insects and raise awareness about the impacts of invasive species. Rather than focusing solely on invasive species, the overlap between the two events got me thinking more broadly about how warming temperatures are changing the composition of Britain’s invertebrate fauna.
I was in London for most of the week (which also coincided with a mini heatwave), taking care of other commitments that unfortunately kept me out of the field – but perhaps that was for the best. With three consecutive days each setting a new June temperature record, the Met Office issued Red Extreme Heat Warnings across parts of England and Wales, warning that ‘hot spells will become more frequent in our future climate’.
You may ask yourself why I am going on about the heat? Well, insects are ectothermic (‘cold-blooded’) so tend to become more active in warmer temperatures, which is why we generally see higher insect activity – and for many groups greater abundance – during summer in temperate zones. However, thresholds of heat tolerance vary across insect groups and species. Bumblebee colonies, for example, are negatively affected by exposure to heat stress through effects on fertility, brood survival rates, and morphology, which then affect forage success rates in workers. But in other species, increased temperatures can be considered a boon. Insects and other invertebrates that can tolerate (or thrive in) extended periods of exposure to high temperatures could become established in areas where they were previously unable to live, such as mosquitoes being recorded in Iceland for the first time in 2025.
These northward (and sometimes altitudinal) expansions or range shifts can be seen across many invertebrate groups. Even Glasgow is seeing new species arrive, either as native species shift their ranges northwards or as non-native species find conditions suitable for establishment.
The Blue Mason Bee (Osmia caerulescens) is one such recent arrival. More commonly found in southern England, there have been sightings of this species in Scotland, including records from Glasgow.
Blue Mason Bee (Osmia caerulescens)
When I first arrived in Glasgow a few months ago, I spent a good deal of time in the Arboretum attached to the Botanic Garden. And as the weather was largely unfavourable for most insects, I found myself flipping stones and bits of dead wood to see what might be living underneath. On one such outing, I discovered three species of non-native flatworm: Kontikia andersoni, the New Zealand Flatworm (Arthurdendyus triangulatus), and the Yellow-striped Flatworm (Caenoplana variegata). All of these species originate from Australasia and have most likely been introduced to the UK through the horticultural trade. The New Zealand flatworm was first recorded in the UK at Edinburgh Botanic Garden in 1950 and is now fairly widespread across Scotland. The other two species are more recent Scottish arrivals; the Yellow-striped flatworm was first found in the UK in 2008 and was initially confined to the south coast of England. Kontikia andersoni was only known from Cornwall and the Isles of Scilly, the Isle of Man, Northern Ireland and the Irish Republic before being recorded in Scotland in 2014. Though these aren’t insects, they illustrate how invertebrate communities can change relative to global trade and changing climatic conditions.
Not far from the Arboretum, I also visited the Botanic Garden glasshouses, where I found two species of non-native ants happily living amongst the tropical vegetation. Linepithema iniquum is an arboreal species native to Central and South America, whilst the Little Yellow Ant (Plagiolepis alluaudi), native to East Africa, has, as recently as 2023, been found living outdoors in continental Europe for the first time. Invasive ants are considered a severe threat to biodiversity, with a recent paper reporting that, on average, 50% fewer individual animals and species are found in ant-invaded areas. As yet, these ants aren’t found outside the glasshouses, presumably because Scottish climatic conditions aren’t currently suitable to support such a move.
Linepithema iniquumLittle Yellow Ant (Plagiolepis alluaudi)Indian Stick Insect (Carausius morosus)
The Kibble Palace glasshouse at the Botanic Garden is also home to a population of Indian Stick Insects (Carausius morosus), which are said to be the descendants of unwanted pets that were released into a ‘good home’ about 20 years ago. Like the ants, they are non-native but are not currently considered invasive in the UK because they have not established outdoor populations. However, it is conceivable that this could happen as the climate warms.
The outlook for invertebrates in a warming climate is a mixed bag with both ‘winners’ and ‘losers’. Some species will likely disappear from areas, and new species will arrive. But this is a complex issue, with multiple additional factors coming to bear on species distributions such as habitat availability, genetic diversity, and species interactions, to name but a few. We can’t be absolutely sure what the impacts of these changes will be, but the scale and pace of them are unprecedented and therefore of concern. Will Indian Stick Insects and other so-called ‘hothouse aliens’ become the new invertebrate fauna of Glasgow as native species experience changing environmental conditions and interact with an increasing number of non-native species? Probably not entirely, but they may one day form part of a significantly altered invertebrate fauna for the city. Many of these newcomers will never become invasive or cause measurable ecological harm, but they nevertheless illustrate how rapidly invertebrate communities are changing in response to climate and human activity.
It has been a whirlwind few months. In March 2026, I moved from London to Glasgow to take up a new role at Glasgow Museums as Collections Officer (Biological Data). I’ve been very excited about taking on the role where I am now responsible for managing the Glasgow Museums Biological Records Centre, through collating, validating, and verifying biological data for the Glasgow City Region (covering 8 local authorities).
This also means that I will be working closely with data providers such as recording schemes and societies, Biodiversity Officers, Coutryside Rangers, environmental organisations, and members of the public to keep the database up-to-date. With over1.3 million records currently held in the database and with growth expected to be around 50,000 new records per year, this is no mean feat. For my first few months in post, I have been familiarising myself with the database and getting to grips with not only a new city and a new country, but also new software systems. It has been a very steep learning curve, and I am very grateful to everyone who has helped out with Recorder 6 queries, field excursions, and introductions to all the right people.
Glasgow Museums Resource Centre. My new office space on the outskirts of Glasgow.
Over the last couple of weeks, my focus shifted from database management to prioritising the delivery of commercial enquiries. The previous pricing structure and commercial offer have been updated so that they are now in line with other Local Environmental Records Centres. This phase of the job is all customer-focused. I have been running some explorative searches from the database and will be meeting with multiple stakeholders to discuss how these data can be used for planning, conservation, research, and public engagement.
In relooking at the offer, I also realised that some of the workflows could be streamlined through using the R programming language and have been working on coding scripts for reporting functions and managing queries.
When not at my desk and behind a screen, I have been getting out to visit green spaces in and around Glasgow with Richard Sutcliffe’s excellent (though sadly now a tad out of date) guidebook as reference. I have only scratched the surface of all the places to visit, but I have been struck by the clear ecological differences between the Scottish lowlands and Southeast England. The climate is obviously cooler and wetter here, and with very different geology as well, come different habitats and therefore different invertebrate species and assemblages.
Loch Lomond near BalmahaRiver KelvinView from Hamiltonhill Claypits LNR
I’m still figuring all this out, but it does mean that I can fairly easily find new lifers when I’m out in the field (or on a lunch break). And, as climate change effects are causing Northward range shifts in invertebrate species, it is also possible to find species newly arrived in the area.
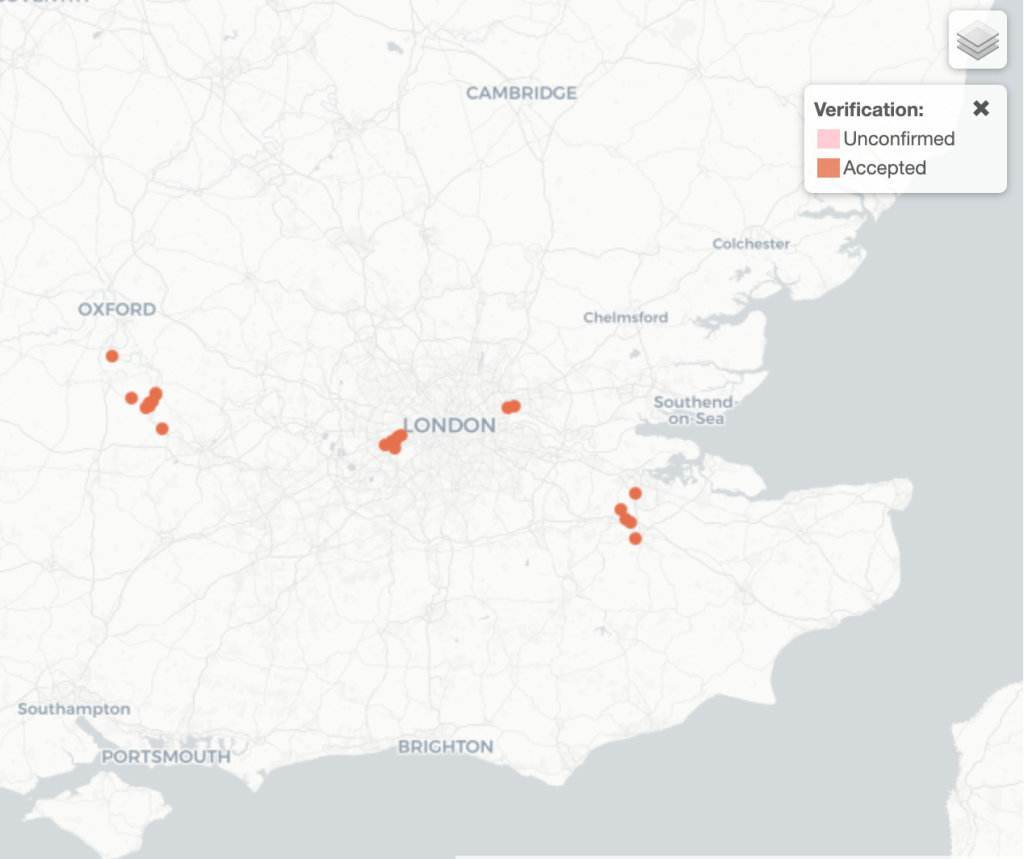
The German Hairy Snail (Pseudotrichia rubiginosa) is a land snail which tends to be found on flotsam along rivers. In the UK, the snail is considered nationally rare and near threatened, being known only from sites along the Thames in Oxfordshire, the tidal Thames in West London, the Roding, and the Medway.
Sites of confirmed records of Pseudotrichia rubiginosa on the NBN Atlas map (Last accessed 23 November 2025)
Hearing the descriptions of habitats where the snail had been found, in both recent and historical sites, I decided that a couple of places near Cody Dock might serve as interesting survey sites. The first, which I thought most likely, was along the banks of what remains of the Channelsea River. The Northern bank of this tidal stretch of water has some fantastic Crack Willow (Salix fragilis) trees, and as such, dead wood and leaf litter tend to collect at the high tide mark. The other site, which seemed more optimistic, was behind the reed bed on the tidal Lea just outside Cody Dock, where scrubbing over had been ongoing for some time.
Once all the partners had been approached, a training day was organised at Syon House – a site in Brentford on the Thames where the snail was known to be found. On the North bank of the Thames, the property was kept as a grazing marsh with many Poplars, Willows, and Swamp Cypress amongst the drainage ditches, an area which the groundskeeper described as “the result of 500 years of benign aristocratic neglect”. Here, we got the opportunity to learn more about the life history of the snail and to trial and hone the survey methodology.
This way for the German Hairy Snail enthusiasts!Syon HouseFinding GHS aplenty
The Conchological Society prepared a handy identification guide to German Hairy Snails (and some lookalikes to be aware of), which can be found here. It was a brilliant day, and every participant found a few of the target snail species, as well as a selection of others, which, if not identified in the field, went off to be confirmed by Dr Ben Rowson from the National Museum of Wales. We left Syon House optimistic that we might find these snails in our survey area near Cody Dock.
With volunteers in wellies and hi-vis vests, we then set out to survey the two sites that we had previously considered. At the first site, on Channelsea, we almost immediately found German Hairy Snails in our first 10m2 plot. We then surveyed another 4 plots and found the snails in every one of them, with a grand total of 97 recorded at the site! However, our second site, behind the reeds on the tidal Lea, did not produce any German Hairy Snail records.
Our first GHS!Many more GHSVolunteers in action – sifting through the debris and plant matter along the high-tide mark at Channelsea River.
It was wonderful to be involved with this project. Many thanks to everyone who helped with the project design and coordination, and to all of our eager volunteers who made the days out in the field so much fun. I am delighted that we were able to add a new site to the distribution maps for this species and hope that these records can support further conservation action along the Channelsea River. I am also very much looking forward to seeing the results from all the other partners involved in the search for this adorable little creature.
Update 24 November 2025:The Guardian newspaper has run an article on the German Hairy Snail project. Follow this link to read the piece.
Further update 24 November 2025:The BBC news website has also run an article on the German Hairy Snail project. Follow this link to read the piece.
It’s been 9 months since I started my role as Biodiversity Officer at Cody Dock and it’s been a rather busy time. I wanted to review some of the work that has been done here, summarising parts of the excellent Tidal Lea Ecology Report prepared by my predecessors & adding what we’ve already managed to build on this with our Community Science and Conservation Volunteers in a concise manner.
From February 2023, I will be taking on the role of Biodiversity Officer for the Gasworks Dock Partnership charity based at Cody Dock on the tidal stretch of the River Lea in east London. I will be co-ordinating volunteer opportunities for surveying and monitoring habitats and wildlife of the local area and delivering related training to our network of community scientists.
Not exclusively focused on invertebrates (though you can be guaranteed they will feature heavily) the monitoring will continue the existing bird counts, bat walks, and vegetation surveys that have been carried out so far – there is an excellent report available that covers this for 2021-2022. Plans are afoot to add in Flower-Insect Timed Counts to contribute to the UK Pollinator Monitoring Scheme and various other invertebrate recording schemes.
Habitat management and improvement works will also be part of this job where establishing new reed beds in the Lea will create cover for water birds and serve as a natural filter for some of the litter and pollutants in the river. Silt traps have already been set along the sloped concrete banks and have become vegetated, and we hope to be able to extend this work further along the east bank. Additionally, there are other areas on or near the industrial estate that can be better managed for wildlife and people including woodland, scrub, and parkland which will entail a number of different projects.
Canary Wharf as seen from the Cody Wilds walk along the east bank of the River Lea.
My vision for this stretch of the Lea is that it acts as a green and blue corridor through this part of east London which has a very industrial heritage, but which is now rapidly being redeveloped with high-density housing. The river serves as the boundary between the boroughs of Newham and Tower Hamlets. These are already densely populated areas (Tower Hamlets has the highest density per km of all English districts, while Newham has the 4th highest population of all the London boroughs) with high levels of poverty (Tower Hamlets has the highest poverty rates in London with Newham ranking 3rd highest). Consulting with property developers at sites in both boroughs to offset some of the habitat loss caused by building apartment blocks as well as helping to shape a nature-friendly approach to the landscaping will also be part of the job. The loss of post-industrial brownfield sites to development is of concern and we will be working to try to mitigate this through the establishment of green roof systems that mimic traditional brownfield habitat as well as advising on the best use of pocket parks and identifying areas to be set aside as wilder habitats.
Bromley-by-Bow Gasworks which is a classic example of the open mosaic habitats of post-industrial brownfield sites in the area which are being redeveloped into high-density housing.
In other news, I recently took on the role of Woodlouse Recorder for the London Natural History Society with plans to increase records for this group so that we can have a better understanding of their distribution across London. So come along to Invertebrate Field Recorder Days across London and Invertebrate Study Days at the Natural History Museum to learn more about these fascinating crustaceans. I have also recently been elected as a trustee for Bethnal Green Nature Reserve where I will be focused on helping with community-led ecology projects.
Working as FSC BioLinks Project Officer has been a fantastic experience. Some of my highlights over the past year-and-a-bit have been my reintroduction to aquatic invertebrates after spending such a long time focussing on all things terrestrial – there is so much to see underwater, and you get to have a bit of splash about which is especially fun on a hot Summer’s day. Formalising my self-taught ant ID with a number of courses (some of which I even got to teach!) and running my version of an Ant Picnic at Richmond Park where I got youngsters to do science while looking at ants. Rediscovering woodlice, millipedes and centipedes; finding the Downland Villa Bee-fly, Villa cingulata, in abundance at Bushy Park; visiting so many amazing sites in and around London from hidden gems to publicly accessible thoroughfares – the list goes on and on.
Downland Villa Bee-fly (Villa cingulata) from Bushy Park in the summer of 2022.
It has been fun and an absolute privilege to work across so many different invertebrate taxa. I recently presented some of the findings from the project at the BioLinks Legacy Conference (the final report will be made available to the public in due course) at the Wellcome Collection in London and am very proud of the work that our team managed to accomplish despite a global pandemic in the middle of our project delivery.
Slide from my presentation at the BioLinks Legacy Conference on 20 January 2023 showing the breakdown of all BioLinks place-based course delivery by invertebrate taxonomic group.
I have learned so much more about a wide variety of invertebrates from national experts (further improving my ID skills) and came to meet a community of people who are passionate, enthusiastic, generous, and knowledgeable about our natural world and the invertebrates upon which we all rely. My sincere thanks to every person who I’ve met along the way and I very much look forward to seeing and working with many of them again in the future.
Set a couple of blocks back from a busy inner city A-road and tucked up against the back of a Sixth Form School is a little patch of greenspace that is the Bethnal Green Nature Reserve. Often overlooked, this volunteer run space offers a little patch of tranquility in the heart of Bethnal Green. I have volunteered here on an ad hoc basis over the year helping with woodland management, pond restoration, turning compost, and whatever else needs attention. This has been a great way to spend a Saturday morning – getting my hands dirty and chatting with other local people who have a stake in the space.
It has also been a perfect place for me to explore my growing interest in invertebrate macro photography and I’m keen to revisit the site again in early 2023 to see what else I can unearth there. There was very little knowledge of the invertebrates living at and using this site with no records submitted to the local environmental records centre. Without any formal recording plan and following the site’s ethos of ‘tread lightly & do no harm’ I have now added 152 invertebrate records across 82 species for the site. And I’m sure this has only scratched the surface.
I was also lucky enough to be invited to spend a few days with some of the site volunteers looking at the different habitat types and the invertebrate assemblage types that are found here. Below are photos taken by the volunteers at these various events.
A series of half-day-long events were organised to explore the invertebrates of Bethnal Green Nature Reserve. Attendance was fantastic and I would like to thank everyone who came along and participated – even those who were a little less keen on our invertebrate neighbours than some others. We looked at nocturnal, pollinating, pond, and leaf litter & soil-dwelling invertebrates. I certainly had a fantastic time and I believe that the volunteers now all have a greater consideration and appreciation for the invertebrate life that is found here. I think this is exemplified by the video below of a Willow Emerald Damselfly (Chalcolestes viridis) captured from the edge of the pond, which was shared in a WhatsApp group by one of the volunteers.
This is a good start. There’s a lot more to be done here in terms of understanding the invertebrate fauna of the site, but there is a willingness, even an eagerness, to do so. I hope that I will be able to support and attend more of these activities over the coming years while we get to know this little urban oasis and all of its many inhabitants better.
To help with this I have now created an iRecord activity where future records from the site can be entered so that they are all kept together and start to build a clearer impression of all the life here as seen and recorded by the people who love and use the site.
I recently wrote a blog post for the FSC BioLinks project on some of the colony founding techniques and behavioural adaptations that have evolved in some ant groups and species. The blog post was entitled The Fantastical Lives of Ants: Slavers, Rebels, and Regicide, but I think I prefer the inversion of the title and subtitle as used above.
The piece was written in response to a comment made by an attendee at one of the Learn to Love Ants courses that I was teaching, where the person remarked that the lives of ants seem much like the fantasy series Game of Thrones. And there are certainly parallels that can be drawn between some of the observed behaviours of ants and the tropes of deception, brutality, conquest, and conflict that are rife in the fantasy genre.
There is of course much more complexity in these social insect societies than just these cherry-picked sensationalist topics, such as: brood care; mutualistic interactions with other organsims; the recently documented care for injured Matabele ants by their sisters; and many more besides. After all, as Sansa Stark says: “I’m sure cutting off heads is very satisfying, but that’s not the way you get people to work together”.
In the run up to both COP 26 and COP 15 many newspapers recently reported the shocking fact that Britain has lost almost half (47%) of its biodiversity since the industrial revolution. For naturalists and conservationists working in the UK this will,however, come as absolutely no surprise whatsoever.
Estimated Biodiversity Intactness Index (BII) in the year 2020 at 0.25 degree resolution. Only the darkest areas have retained enough natural biodiversity to be within the proposed planetary boundary (where BII is above 90%).
Research by Prof Andy Purvis from the Natural History Museum in London showed that Britain is one of the most nature-depleted nations in the world, well below the global average of 75%. With the publication of the Biodiversity Intactness Index (BII) we can now clearly see in the data what naturalists have been warning about for decades from their field observations – Britain’s biodiversity is in peril.
What’s the deal with biodiversity anyway?
‘…Biodiveristy provides us with the food we eat, from the micro-organisms that enrich the soil where we grow our crops, to the pollinators who give us fruit and nuts… [and] many of our medicines originate from plants and fungi…’.
Sir Richard Attenborough
This beautiful animation (below) narrated by Sir David Attenborough and produced by The Royal Society explains the importance of biodiversity, both to us and the world at large.
When 67% of the UK is used for agriculture and a further 8% is built on that leaves a paltry and dwindling 25% for nature. According to official statistics from the Ministry of Housing, Communities and Local Government (2018), forest, open land and water constitute 21% of all land use in England.
‘As Presidents of COP26, the UK has put nature at the heart of the agenda, and we very much welcome this important study which highlights the crucial connections between climate and biodiversity and the urgent need to protect nature’.
Lord Zac Goldsmith, UK Government Minister for Pacific & the Environment
Damningly though, researchers from the RSPB have found that although 28% of UK land is reported by the UK government to be protected, only 11.4% of land area actually falls within protected areas designated primarily for nature conservation. And because of the poor condition of some of these areas, as little as 4.9% of UK land area may in reality be effectively protected for nature.
How do we effectively address this issue in Britain?
‘Governments possess the power – economic, political and legal – to address the planetary emergency, and there may still be time, but they must act now.’
Prof Andy Purvis, Natural History Museum
The British Ecological Society produced a report in May this year (2021) that called for a nature-based approach to tackling both climate change and biodiversity loss in conjunction with other climate and conservation actions. A brief summary of their specific policy recommendations provide examples of opportunities across a range of habitats through:
Restoring degraded peatlands and end burning on blanket bogs
Increasing native woodland and woodland connectivity in the right places
Establishing more saltmarshes
Protecting and re-establishing hedegrows in arable landscapes
Increasing agroforestry in arable landscapes
Increasing urban green spaces with a focus on native species
Unfortunately, any and all action to prevent further biodiversity loss is costly. A recent report from the Green Finance Institute claims that the UK governement faces as much as a £97 billion funding gap for its current commitments to nature-based actions over the next 10 years.
Regardless of the financial costs of mitigating and remedying biodiversity loss, we should never lose sight of the costs of inaction – not just economic, though these are significant. But also the legacy of a pillaged, spoiled and empty landscape; a depauperate and diminished native biota; and ultimately, an impoverished and increasingly precarious society.
Instead of attending an in-person seminar this year, PhD students in our department at UCL were recently asked to produce a video in response to a question set by the Post-graduate Tutors.
This is my video responding to the question: “What have I learned (so far) during my PhD?”.