From February 2023, I will be taking on the role of Biodiversity Officer for the Gasworks Dock Partnership charity based at Cody Dock on the tidal stretch of the River Lea in east London. I will be co-ordinating volunteer opportunities for surveying and monitoring habitats and wildlife of the local area and delivering related training to our network of community scientists.
Not exclusively focused on invertebrates (though you can be guaranteed they will feature heavily) the monitoring will continue the existing bird counts, bat walks, and vegetation surveys that have been carried out so far – there is an excellent report available that covers this for 2021-2022. Plans are afoot to add in Flower-Insect Timed Counts to contribute to the UK Pollinator Monitoring Scheme and various other invertebrate recording schemes.
Habitat management and improvement works will also be part of this job where establishing new reed beds in the Lea will create cover for water birds and serve as a natural filter for some of the litter and pollutants in the river. Silt traps have already been set along the sloped concrete banks and have become vegetated, and we hope to be able to extend this work further along the east bank. Additionally, there are other areas on or near the industrial estate that can be better managed for wildlife and people including woodland, scrub, and parkland which will entail a number of different projects.
Canary Wharf as seen from the Cody Wilds walk along the east bank of the River Lea.
My vision for this stretch of the Lea is that it acts as a green and blue corridor through this part of east London which has a very industrial heritage, but which is now rapidly being redeveloped with high-density housing. The river serves as the boundary between the boroughs of Newham and Tower Hamlets. These are already densely populated areas (Tower Hamlets has the highest density per km of all English districts, while Newham has the 4th highest population of all the London boroughs) with high levels of poverty (Tower Hamlets has the highest poverty rates in London with Newham ranking 3rd highest). Consulting with property developers at sites in both boroughs to offset some of the habitat loss caused by building apartment blocks as well as helping to shape a nature-friendly approach to the landscaping will also be part of the job. The loss of post-industrial brownfield sites to development is of concern and we will be working to try to mitigate this through the establishment of green roof systems that mimic traditional brownfield habitat as well as advising on the best use of pocket parks and identifying areas to be set aside as wilder habitats.
Bromley-by-Bow Gasworks which is a classic example of the open mosaic habitats of post-industrial brownfield sites in the area which are being redeveloped into high-density housing.
In other news, I recently took on the role of Woodlouse Recorder for the London Natural History Society with plans to increase records for this group so that we can have a better understanding of their distribution across London. So come along to Invertebrate Field Recorder Days across London and Invertebrate Study Days at the Natural History Museum to learn more about these fascinating crustaceans. I have also recently been elected as a trustee for Bethnal Green Nature Reserve where I will be focused on helping with community-led ecology projects.
Working as FSC BioLinks Project Officer has been a fantastic experience. Some of my highlights over the past year-and-a-bit have been my reintroduction to aquatic invertebrates after spending such a long time focussing on all things terrestrial – there is so much to see underwater, and you get to have a bit of splash about which is especially fun on a hot Summer’s day. Formalising my self-taught ant ID with a number of courses (some of which I even got to teach!) and running my version of an Ant Picnic at Richmond Park where I got youngsters to do science while looking at ants. Rediscovering woodlice, millipedes and centipedes; finding the Downland Villa Bee-fly, Villa cingulata, in abundance at Bushy Park; visiting so many amazing sites in and around London from hidden gems to publicly accessible thoroughfares – the list goes on and on.
Downland Villa Bee-fly (Villa cingulata) from Bushy Park in the summer of 2022.
It has been fun and an absolute privilege to work across so many different invertebrate taxa. I recently presented some of the findings from the project at the BioLinks Legacy Conference (the final report will be made available to the public in due course) at the Wellcome Collection in London and am very proud of the work that our team managed to accomplish despite a global pandemic in the middle of our project delivery.
Slide from my presentation at the BioLinks Legacy Conference on 20 January 2023 showing the breakdown of all BioLinks place-based course delivery by invertebrate taxonomic group.
I have learned so much more about a wide variety of invertebrates from national experts (further improving my ID skills) and came to meet a community of people who are passionate, enthusiastic, generous, and knowledgeable about our natural world and the invertebrates upon which we all rely. My sincere thanks to every person who I’ve met along the way and I very much look forward to seeing and working with many of them again in the future.
Set a couple of blocks back from a busy inner city A-road and tucked up against the back of a Sixth Form School is a little patch of greenspace that is the Bethnal Green Nature Reserve. Often overlooked, this volunteer run space offers a little patch of tranquility in the heart of Bethnal Green. I have volunteered here on an ad hoc basis over the year helping with woodland management, pond restoration, turning compost, and whatever else needs attention. This has been a great way to spend a Saturday morning – getting my hands dirty and chatting with other local people who have a stake in the space.
It has also been a perfect place for me to explore my growing interest in invertebrate macro photography and I’m keen to revisit the site again in early 2023 to see what else I can unearth there. There was very little knowledge of the invertebrates living at and using this site with no records submitted to the local environmental records centre. Without any formal recording plan and following the site’s ethos of ‘tread lightly & do no harm’ I have now added 152 invertebrate records across 82 species for the site. And I’m sure this has only scratched the surface.
I was also lucky enough to be invited to spend a few days with some of the site volunteers looking at the different habitat types and the invertebrate assemblage types that are found here. Below are photos taken by the volunteers at these various events.
A series of half-day-long events were organised to explore the invertebrates of Bethnal Green Nature Reserve. Attendance was fantastic and I would like to thank everyone who came along and participated – even those who were a little less keen on our invertebrate neighbours than some others. We looked at nocturnal, pollinating, pond, and leaf litter & soil-dwelling invertebrates. I certainly had a fantastic time and I believe that the volunteers now all have a greater consideration and appreciation for the invertebrate life that is found here. I think this is exemplified by the video below of a Willow Emerald Damselfly (Chalcolestes viridis) captured from the edge of the pond, which was shared in a WhatsApp group by one of the volunteers.
This is a good start. There’s a lot more to be done here in terms of understanding the invertebrate fauna of the site, but there is a willingness, even an eagerness, to do so. I hope that I will be able to support and attend more of these activities over the coming years while we get to know this little urban oasis and all of its many inhabitants better.
To help with this I have now created an iRecord activity where future records from the site can be entered so that they are all kept together and start to build a clearer impression of all the life here as seen and recorded by the people who love and use the site.
I am a self-professed invertophile. I absolutely adore the myriad forms of insects and other spineless creatures. They are the most diverse and abundant group of organisms on earth, they can be found in every habitat imaginable, they have evolved some of the most complex forms, lifestyles and behaviours, and they are responsible for maintaining essential ecosystem functions and systems. How could one not be utterly awed by them?
Nemopoda nitidula an ant-mimicking fly from the ensign fly family Sepsidae.
I grew up in South Africa and lived on the outskirts of a small town in KwaZulu-Natal. I played on the edge of wilderness and ‘civilisation’ where the veldt and acacia scrub met our mowed lawn and meticulously weeded flower borders. The garden was surrounded by a low wall built from great chunks of blue-grey and rust-coloured igneous rocks (which were displaced by the flower beds) and poured concrete. I travelled in a circuit along these walls and around the garden marvelling at all the life that was to be found here: Citrus Swallowtail butterflies and their peculiarly pungent caterpillars that were resident in our lemon tree; ants that magically appeared around every dropped crumb; checkered yellow and black blister beetles which I knew not to touch; the iridescent snap of a dragonfly’s wings as it hawked overhead. I also kept my share of ‘pets’ that wandered too close to the house and ended up living in jam jars with holes punched through the lid. As I grew older the farther I wandered from the borders described by the walls, drawn further and further away by the towering curiosities that rose out of the earth and teemed with thousands of milky-white termites. I watched trapdoor spiders snatch up prey, ran with solifugids and scampered from scorpions. I carefully turned over logs and rocks and watched centipedes and beetles scurry from the light. I listened to the susurrus hiss of grasshoppers, and when those turned to the chirps of crickets I knew that it was time to head home for dinner. My childhood summers were glorious and almost every day was filled with LIFE.
The Painted Lady (Vanessa cardui) is a long-distance migrant with each journey from tropical Africa to Europe taking up to six generations to complete.
Now, much later in life and living in London, I still find the presence of wild animals very rewarding. And still, none more so than the invertebrates. They are perhaps not as abundant or as large as those of my youth, but they are all around us even if we need to look a little harder. I now have a number of local patches where I go to observe invertebrates. All within a comfortable walking distance of my apartment and all quite different from one another: a local park, a cemetery and urban nature reserve, a city farm, a medicine garden and community space, and a brownfield site. So far this year I have recorded many species new to these sites, several new to the borough and new to me!
Although I understand that not everyone shares my passion for the myriad creatures that surround us and that some people can be downright hostile towards them; I can’t help but feel that they’re missing out on something quite incredible. And to that point I’ve been thinking a lot about observing and recording invertebrates recently – specifically about how people might get started with it.
How to get involved
There are many ways in which you can become involved with observing, identifying and recording invertebrates. Here, I will specifically discuss casual recording – by this I mean randomly walking through a space of your choosing and observing invertebrates in situ. There are no formalised procedures, no sampling methodologies, just you in nature. I think that this is a great way to become familiar with the variety of life out there. Having said that, you will find a few items incredibly useful for helping you along your new voyage of insect discovery:
Comfortable walking shoes,
A good introductory or general field guide,
A camera (a phone camera will most often work well enough),
A GPS or phone that can give you location coordinates,
A notebook and pen/pencil,
A 10x magnification hand lens.
I will follow this blog post up with another about different invertebrate sampling methods in the future.
A note on some of the field guides that are available: the Collins Complete Guide to British Insects by Michael Chinery, though by no means complete, is a decent place to start as it covers many of the more common species (>1,500) and was in fact my first field guide. I then moved on to Paul Brock’s A Comprehensive Guide to Insects of Britain and Ireland which, though not comprehensive, goes somewhat further than Chinery, covering more species (2,300) and also includes some of the rarer insects. Most recently (2021) Brock has published Britain’s Insects with WILDGuides which focuses on more popular groups and species. This is an interesting publication with some excellent entries and photography, but covers a reduced number of species (1,653). I own and regularly use all of these but also more specialist guides to various groups of insects. However, when starting out, I would recommend that you get one of these to use in the field and will point out some of the excellent online resources and forums that are also available. Please note that these books are available through other bookshops and online sellers, I have linked to NHBS as they actively support conservation.
The mayfly, Ephemera vulgata, though common throughout Europe is in decline. It is thought that this may be due to the pollution of waterways.
How to find invertebrates
Stop. Pick a spot and stand still.
Get your eye in. Let your eyes slowly scan across the vegetation in front of you just below eye-level. Look for movement, see if there are any odd shapes or colours that stand out from the background. Remember many insects can be very well camouflaged so take your time.
Get down low. I tend to crouch a lot, but you could also kneel or sit on the ground. If you’re low down you will be more likely to see ground-dwelling invertebrates. This is why young children make fantastic “bug hunters”.
Listen. Some insects will make noise to attract mates like crickets and grasshoppers, but you can also hear the snap of dragonfly wings, the rustle of grass as something moves through it, and even the munching of leaves.
Move slowly and carefully. Don’t move far, but move a few steps at a time while keeping an eye on where you place your feet. As you move you want to try to avoid disturbing the vegetation as much as possible as invertebrates can be very sensitive to vibrations. Also, beware your shadow as this can frighten off the flightier individuals.
Look forsigns of invertebrate presence. Nibbled leaves, cut stems, silk threads, nest holes and the like. Sometimes even tracks in sand can be signs that invertebrates are about; and always keep an eye out for frass (essentially larval poop).
Don’t forget to look up. Remember that many insects can fly. Also, it is definitely worth examining vegetation at or just above head height.
Make notes and/or take photos. This is very useful for your own future reference, but also if you want to report your sightings to any of the recording schemes. I will talk about this in a bit more detail later on, but basic information that is useful is: a photo, the date, species name, number seen, and location.
Through recording wildlife we can determine a number of important data about what animals are found in which habitats. With long-term data we can see if these species change over time and this can help us to understand the drivers of those changes e.g. habitat loss, pollution events, land restoration etc. We can track the movement of species’ distributions in response to large-scale and seasonal effects such as climate change, and we can monitor the conservation status of species in order to identify those most at risk of extinction. Invertebrates are specifically important because it is in their changes that we tend to first detect issues of future conservation concern. I hope that I’ve managed to convince you that this is a worthwhile project to undertake for better understanding these incredible creatures that share the planet with us.
In Britain the recording community is largely voluntary, from people going out into the field to record what’s in their local patch to the experts who verify these records and the county or national recorders who collate it all. There are of course exceptions such as ecologists who might be employed to survey sites for invasive species or for endangered species that might affect construction projects. But for the most part people survey and submit records for their own personal reasons which can be as varied as the number of people involved; whether that’s about wanting to contribute to scientific enquiry, wanting to know more about the wildlife in a local area, or wanting to catch them all…
Lasius brunneus, the Brown Tree Ant has only been recorded from central and southern English counties despite suitable habitat being available across Britain. It is thought that its arboreal and timid nature mean that it is often overlooked and therefore under-recorded.
How to record invertebrates
Recording invertebrates is a two-step process. The first step is what information is kept in your field notebook. I tend to record a bit more information here than I will need for submitting to the recorders/recording societies.
On a new page in my field notebook I always start with this information:
Date
Weather – the general outlook for the day.
Site name
Site notes – you may want to specify habitat type(s) or whether there has been any site management or disturbance since your last visit etc.
Casual recording – or specify which sampling method was used.
I then start searching for invertebrates and record them each like this:
Species name – if known, otherwise genus or family and update it later.
Male / Female / Mixed – if you can tell, it isn’t always possible.
Life stage – adult, larva, nymph, pupa, etc.
Identified by – this is if someone else has helped you with an ID.
Number – you need to decide on the scale you want to use here, I tend to include all individuals within 102 metres, but you can extend this to 1002 m or 12 km if you want to include a whole site.
Coordinates – I normally get latitude and longitude from my phone using either Google Maps or Apple Maps in decimal format.
Photo number – if using a camera that records this information.
Notes – any significant interactions or interesting behaviours.
And that’s it!
The second step is to submit your records and there are a few different ways in which you can do this. For the most part I use the online recording website iRecord which a large number of verifiers and recorders use. For more information about how iRecord works take a look at this blog post and video produced by Keiron Derek Brown.
Alternatively you can manage your own database in Excel and provide these records to the national recording scheme or relevant recorder directly via email if that’s what they would prefer.
This is the first observation of the endangered picture-winged fly Tephritis praecox in Tower Hamlets and only the second record in Middlesex since 2016.
The more you look…
I have lived in Tower Hamlets for 10 years and in the last few months I have been incredibly fortunate to find three endangered insect species in some of my local patches. This is because I have spent more time looking and got lucky. This is what makes casual recording so exciting for me, you just never know what might turn up.
Oberea oculata, the critically endangered and very scarce, Eyed Longhorn Beetle. I was able to add a new site to the previously known distribution of this species.
The title of this blogpost is taken from the latest BBC television series that has just finished screening in the UK. The premise for this series is based on the model developed for the hugely successful The Great British Bake Off, in which contestants compete against each other day-after-day to produce a variety of baked goods that are then judged by experts. Replace baked goods for ceramics and you will have grasped the intricacies of The Great Pottery Throw Down in its entirety. But what, if anything, is the significance of this to the study of invertebrates?
Well, having recently read Animal Architecture by Ingo Arndt and marvelled at the complexity and ingenuity of animals to create structures such as the heaped nests of wood ants, the towering cathedrals of termitesand the delicately partitioned nests of paper wasps; I was rather taken with the notion of insects as ‘makers’. Serendipitously, I stumbled across the website of naturalist and artist, John Walters– specifically across his marvellous illustrations and accounts of Heath Potter Wasps, Eumenes coarctatus.
Eumenes coarctatus. Source: Wikipedia
Potter wasps, Eumeninae, are the most diverse group of Vespidae, with over 3,500 species in 210 genera found throughout the world. Of these, 23 species in 9 genera are found in the UK. They derive their common name of potter (or mason) wasps from the fact that the females tend to construct nests from mud and clay. These nests can take multiple forms, but one of the most elegant (in my view) is the vase-shaped nest of Eumenes coarctatus, the solitary Heath Potter Wasp.
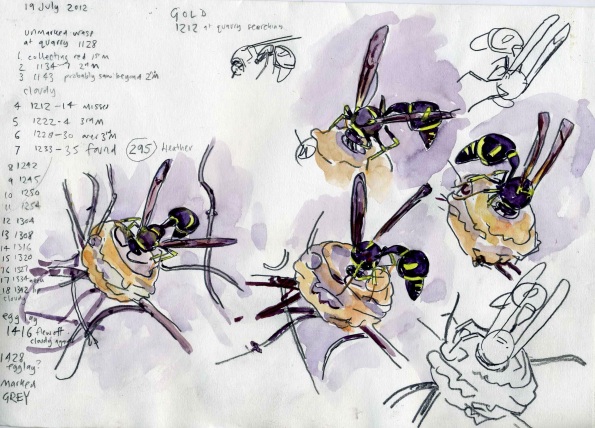
Using heather, gorse or dead grass stems as nesting sites, the female will build her clay vessel over the course of two to three hours. During this time she will repeatedly fly from a water source to a quarry site, where she will form a ball of mud in her jaws, which is then transported to the construction site where she builds the nest. Once she has shaped the neck and lip of the nest she lays a single egg in the chamber suspended on a strand of silk. She will then search for, sting and collect a number of small caterpillars, especially pug and horse chestnut moth larvae, from the heathland vegetation and then fills the pot with them. A final trip to the water source and quarry then provides enough clay to seal the pot with between 9 and 38 paralysed caterpillars trapped inside. A female heath potter wasp may produce up to 25 pots in her lifetime (2 to 3 months) and occasionally she will cluster pots as shown in the series of photographs below. It is possible that these clusters prefigure the development of eusocial colonies as seen in some other vespids.
Development of a cluster of clay nest cells built by a single Eumenes coarctatus. Bovey Heath, Devon. Photos by John Walters.
When the wasp larva hatches from its suspended egg it drops onto the paralysed prey on which it feeds for about a week before pupating. The emergence of the adult depends on the timing of the building of the pot. If the pot was built before the end of June, the adult wasp will emerge 2 to 3 weeks later; if the pot is built in early July, the adult will still emerge in the same year; however, if the pot is built after this date emergence will be delayed until the following April or May.
Illustration of Heath Potter Wasps taken from the field notes of artist and naturalist John Walters.
Though the E. coarctatus larvae are predatory, the adults feed on the nectar of heathland plants such as gorse, heather, bramble, angelica and alder buckthorn.
Distribution map of Eumenes coarctatus in the UK. Source: NBN
Found on lowland heaths in England (from South Devon to East Sussex, and north to Buckinghamshire) the Heath Potter Wasp is classified as nationally scarce, though not designated a BAP species.
I also love Jean-Henri Fabre’s description of Eumenes taken from The Wonders of Instinct:
“A wasp-like garb of motley black and yellow; a slender and graceful figure; wings not spread out flat, when resting, but folded lengthwise in two; the abdomen a sort of chemist’s retort, which swells into a gourd and is fastened to the thorax by a long neck, first distending into a pear, then shrinking to a thread; a leisurely and silent flight; lonely habits.”
More:
John Walters has also made a video of a wasp building a nest.
Michael Archer’s Key to British Potter and Mason Wasps is a very useful resource for identifying the various UK species.
There are more than 500 species of Bombardier beetle (a form of ground beetle – Carbidae) in the tribes Brachinini, Paussini, Ozaenini, or Metriini all displaying the highly effective defence mechanism of releasing a superheated pulsing jet of noxious chemicals sprayed directly at would-be predators. I have always been fascinated by the ammunition of Bombardier beetles – their highly accurate and violent chemical attack brought on whenever you touch them – but it was only on reading Eisner’s essay that I started to fully grasp the incredible complexity of these beetles.
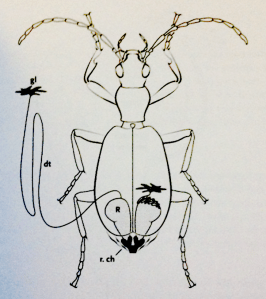
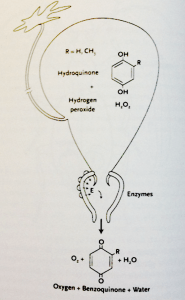
These diagrams are reproduced from Thomas Eisner’s fascinating book For Love of Insectswhich explores the variety of ways in which insects use chemicals for defence, signalling and prey capture. I cannot recommend this book highly enough for anyone interested in the pursuit of entomology or study of chemical ecology.
Diagram of a bombardier beetle with its 2 glands in place. R = reservoir; r.ch = reaction chamber; gl = glandular tissue; dt = duct
The mechanism of operation of the bombardier glands. E = enzymes in the reaction chamber; R indicates that either a hydrogen (H) atom or a methyl group (CH3) can occur at that site on the hydroquinone or quinone molecule.
The chemical process involves hydrogen peroxide rapidly decomposing into oxygen and boiling water, while the hydroquinones oxidize into benzoquinone in the beetle’s reaction chambers. This mix explodes out of the beetle with an audible popping sound, in a volley of rapid-fire blasts – in a manner likened to the pulsing propulsion system of Germany’s V-1 “buzz bomb” in WWII. The consequent foul chemical burn (at 100°C) incapacitates smaller attackers like ants, and deters larger predators such as the unfortunate frogs in one of Eisner’s experiments.
By examining the propulsion mechanism using high-speed synchrotron X-ray imaging Eric Arndt from MIT confirmed Eisman and his colleagues’ qualitative passive ‘pulse jet’ model. This researchshows that a flexible membrane and a valve passively control the spray pulsation – as pressure increases in the reaction chamber because of the chemical explosion, the membrane stretches and the valve closes. The membrane then relaxes and the valve reopens once the pressure has been reduced following the ejection of the liquid, and so the process repeats.
The only UK resident species of bombardiers are Brachinus sclopeta, the streaked bombardier beetle, and Brachinus crepitans, the common bombardier beetle, and both are rarely seen. B. sclopeta is so rare that it has only recently been accepted as a native species (past records were thought to be of rare migrants) and prior to 2005 had been presumed extinct since 1928. Because the beetles prefer habitats with thin soil, rubble and bare ground they tend to favour brownfield sites and have been found in east London; but with the continuous and unrelenting development of this area, these beetles’ futures are very precarious despite being listed as UK BAP priority species, and considered critically endangered by the IUCN. However, insect charity Buglife worked with developers to secure a site near London City Airport that is now the only known intact colony of the species in the UK. You can read Richard Joneshighly informative blog about the relocation and conservation of this species.
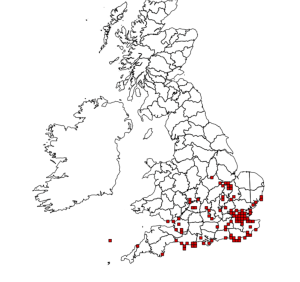
Brachinus crepitans, though more common than B. sclopeta, is restricted to Southern England and Wales and especially the coastal areas of the South-East where it is considered nationally scarce. It is more commonly found in continental Europe, central Asia, the Middle East and North Africa, with central Sweden being the northernmost extreme of its range.
Brachinus crepitans. Image source: Wikipedia
Distribution of Brachinus crepitans. Source: NBN Gateway
Usually seen in May and June, the beetle favours calcareous grasslands, arable field margins and chalk quarries. It is usually found in dry, sunny areas – typically under stones. Little is known about its life-cycle, but it is thought that the larvae are external parasites on the pupae of other species of beetle, particularly those of the ground beetle Amara convexiuscula and a staphylinid beetle, Ocypus ater.
I think these gorgeous and enthralling beetles definitely warrant being surveyed for this coming Summer.
References:
Eisner, T. (2003) For Love of Insects. Harvard University Press, Cambridge Ma.
Lyneborg, L. (1976) Beetles in colour. Blandford Press, Dorset.
The Indian or laboratory stick insect, Carausiusmorosus, a common pet.
The order Phasmatodea contains more than 3,000 extant species of insect found throughout the world, especially the warmer zones. These herbivorous (mostly arboreal) insects are most well-known for their crypsis, or camouflage, where their colour, shape and behaviour enable them to masquerade as twigs or leaves (hence their common names of stick and leaf insects).
Though there is still uncertainty about a definitive phylogeny, phasmids are considered one of 11 orthopteroid insect orders within the assemblage known as Polyneoptera. Phasmatodea are found alongside the modern orders: Blattodea (cockroaches), Dermaptera (earwigs), Embioptera (web-spinners), Grylloblattodea (ice crawlers), Mantodea (praying mantises), Mantophasmatodea (heel-walkers) and, of course, Orthoptera (crickets, grasshoppers, and katydids). According to phylogenomic analyses of nucleotide and amino acid sequences, some of these Polyneopteran lineages are thought to have emerged ~302 million years ago, with phasmids evolving after the Permian mass extinction.1
Fossil Evidence
Fossil evidence of phasmids is, however, extremely rare. Specimens have been recovered in amber, most notably representatives of Euphasmatodea and Timematodea, with the oldest well-documented fossils being found in Cretaceous Burmese amber. Recent discoveries of the oldest-known fossilised leaf mimics (Phylliinae) from Messel, Germany in 2006 of Eophyllium messelensis date this foliacious mimicry to the Eocene.2
Although the fossil evidence is patchy, it is thought that traits relating to morphological plant masquerade within Phasmatodea first developed with stick mimicry in the Permian, followed by leaf mimesis developing in the Eocene when angiosperms largely replaced conifers as dominant trees.2
Simplified cladogram with a partial geochronologic scale showing the phylogenetic position of E. messelensis and the temporal sequence of character evolution. Insert; Photo (A) of holotype of fossil leaf insect E. messelensis, from the Eocene Messel Pit, Germany and Photo (B) Cretophasmomina melanogramma from 126 mya from the Yixian formation in Inner Mongolia [2, 3]Fossils of Cretophasmomima melannogramma discovered in Yixian, China in 2006 from the Cretaceous Jehol biota (approximately 129 mya), however, provides evidence of phasmid crypsis relating to a Gingkophyte, Membranifolia admirabilis, which displayed leaf-shaped plant organs.3 It is therefore thought that leaf mimesis may well have developed far earlier than previously thought.
Crypsis, Camouflage, and Masquerade
In order to avoid visual detection by predatory mammals, birds, reptiles and other invertebrates, many insects evolved morphological characteristics that enabled them to blend in to their surrounding environment. Specifically in the case of phasmids, these evolutionary adaptations have been very closely linked to the insects’ host and food plants leading to a coupling of ecological and evolutionary dynamics.4,5,6
Cryptic colouration, elongation of the body and legs, or, alternatively, broadening and flattening of the body to resemble leaves are all forms of masquerade adopted by phasmids to avoid detection by predators. This has led some researchers to conclude that predation may be an important driver of speciation in this order. Successful adaptation, through camouflage, may therefore lead to divergence in adaptive radiation. 4,5,6
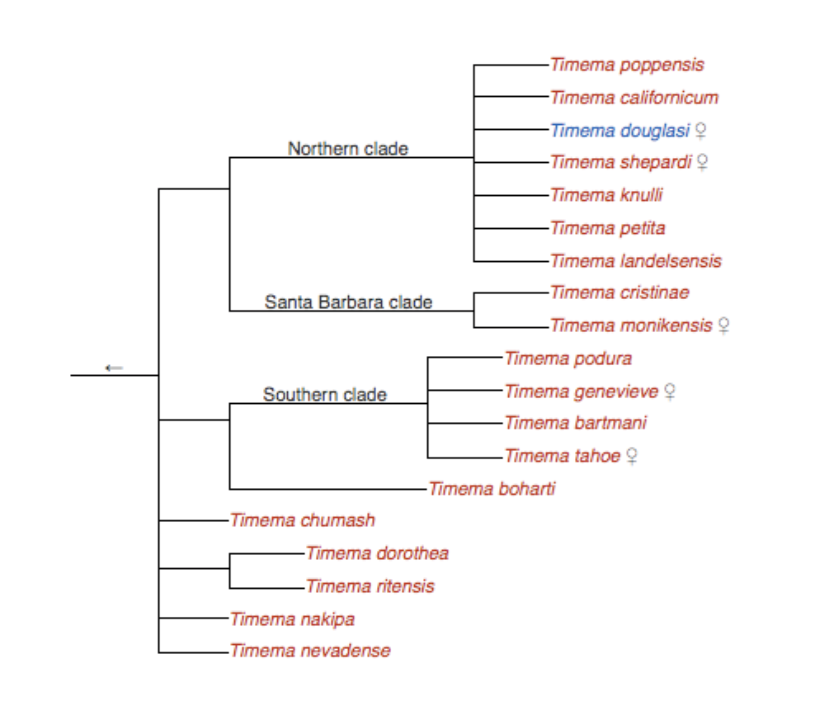
The basal-most extant recorded clade of Phasmatodea is the sub-order Timematodea, within which is found the genus Timema whose species are found throughout southwestern North America on a variety of host-plant species.7 Experimental studies have been conducted into the influence of ecological factors with regards adaptive radiation in Timema cristinae with particular emphasis on host plants and cryptic colouration.4,5,6,7
Timema cristinae, endemic to California has repeatedly evolved ecotypes adapted to different host plant species. One phenotype has no stripe and feeds on Ceanothus (left).
One ecotype features a distinct white stripe on its back and feeds on the thin, needle-like leaves of a shrub called Adenastoma.
Above: Timema cristinae, endemic to California has repeatedly evolved ecotypes adapted to different host plant species. One ecotype features a distinct white stripe (right photo) on its back and feeds on the thin, needle-like leaves of a shrub called Adenastoma. The other phenotype has no stripe and feeds on Ceanothus (left). Below: Phylogeny of Timema species.
It was found that two distinct T. cristinae morphs had developed on two morphologically dissimilar plant species distributed in parapatric mosaics. The first plant, Adenostoma fasciculatum, has needle-like leaves, while the other, Caenothos spinosus, has broad, ovate leaves. Each T. cristinae morph (or ecotype) was found to be more cryptic on one of the two plant species depending on whether they displayed a heritable white dorsal stripe or not. One of the experiments found that bird predation significantly lowered numbers of T. cristinae that were maladapted to the host plant.4 Further studies concluded that through predator pressure, partial (but incomplete) ecological speciation has occurred in T. cristinae as the morphs still successfully interbred.5,6,7
This partial speciation may, however, form only one dimension of selective pressures that constitute adaptive radiation events. It was also shown that by comparing different Timema species that share the same host plant, that sexual isolation was not as marked as with between species on different plants (when compared with T. podura and T. chumash)7 lending credence to the notion of ecological speciation. It is therefore apparent that predators apply selective pressure leading to morphological crypsis and divergence, but that this does not necessarily directly lead to speciation, but is more likely an intermediate stage in adaptive radiation.
Flying, Jumping, and Holding Still
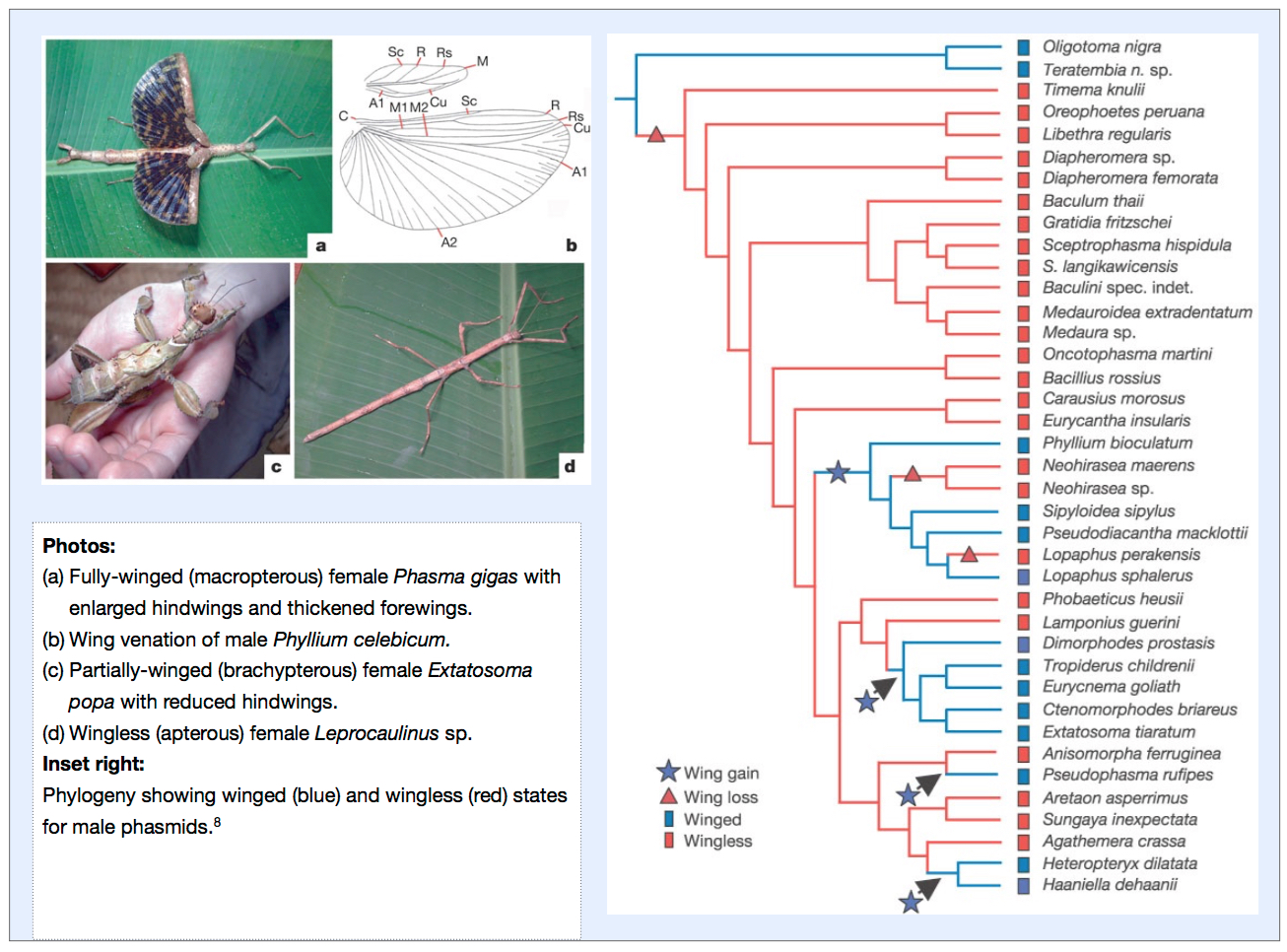
In the case of the Phasmatodea, it has been discovered that diversification occurred in a wingless state and that wings were subsequently derived on a number of occasions.
Of the 3,000 species of phasmids, only 40% are fully winged, while the remainder are partially winged or entirely wingless. While being fully winged conveys advantages of dispersal, escape and finding resources, it has been claimed that increased female fecundity and crypsis may have served as a selective advantage in early phasmid evolution in a shift to winglessness. Apart from flight, wings and partial wings can also be used in threat or startle responses to deter would-be predators. By examining DNA sequence data and applying parsimony optimisation it was found that the ancestral condition of Phasmatodea is wingless.8
It was also found that certain phasmid lineages had “re-evolved” wings prompting suggestions that this reacquisition may confer adaptive advantages of being both winged and wingless as conditions necessitate over ecological time leading to further speciation.8
Timema chumash is unusual in phasmids in that it has been found to jump away from potential threats. Although it can only jump relatively short distances by extending the hind tibia, it can reach take-off velocities comparable to some larger European flea beetles. The leg positions and hind-leg length of T. chumash contrast with the morphology of other stick insects; its legs emerge ventrally from the thorax and its hind legs are proportionately longer than those of other phasmids. As T. chumash is wingless, it jumping is suggestive that it would enable a rapid fall from the plant it was perching on taking it out of the visual field of predatorsand providing it with another opportunity to camouflage itself nearby.9 These morphological and behavioural traits may present opportunities for further adaptive evolution.
A well-documented behaviour in many phasmids is that of catalepsy whereby the insect is able to remain motionless or produce extremely slow movement as a form of twig or leaf mimesis to aid with predator evasion. The mechanism by which this is achieved, is via the high gain of the femur-tibia joint control system,10 and has been recorded in fossil specimens.2 A key difference between phasmids and other orthopteroids is this significant coevolution of the mimetic body shape with catalepsy.10
Parthenogenesis, Hybridogenesis and Androgenesis
Phasmids experience a wide array of reproductive modes with about 10% of the phasmid taxa being parthenogenetic and producing all-female offspring (thelytoky).11 Although parthenogenesis reduces genetic variability, it does not wholly suppress it. Furthermore autopolyploids and allopolyploids can take advantage of higher mutational rates to increase heterozygosity.Androgenesis is also common and has been proposed as a likely pathway to cladogenesis in the genus Clonopsis11and has already been recorded in Pijnackeria where tetraploid hybrids lacking maternal genes, but keeping the maternal mitochondrial DNA, speciated.12 The discovery of interracial and interspecific hybridogenesis in the genus Bacillus added further weight to the notion of maintaining (or even increasing) genetic diversity within phasmid lineages and creating opportunities for further speciation.12
No one reproductive mechanism is exclusively used, so that complete reversion from thelytoky to amphimixis is possible. These “tangled interactions” allow for genetic diversity to persist within and between populations. When considered as part of a series of repeated and complex reproductive strategies including sexual reproduction, parthenogenesis, androgenesis and hybridogenesis, it must be concluded that evolutionary pathways for phasmids are far from dead-ends.11, 12
Following divergence from other orthopteroids, phasmids took advantage of the new food sources and flourished following the angiosperm revolution and have continued to adapt in relation to predatory pressures, host-plant availability, behaviours, and complex reproductive strategies. Clearly, apart from the opportunities presented in the Eocene for cladogenesis and speciation, there continues to be further evolutionary opportunities relating specifically to morphology and sexual isolation in adaptive radiation of phasmids.
The Black Beauty stick insect, Peruphasma schultei, is known to exist only in a tiny area of 5ha (12 acres) in the Cordillera del Condor region of northern Peru, at altitudes between 1200-1800m.
References:
1. Misof, B. et al. (2014), Phylogenomics resolves the timing and pattern of insect evolution. Science. 346 (610), 763-767.
2. Wedmann, S., Bradler, S., and Rust, J. (2006), The first fossil leaf insect: 47 million years of specialized cryptic morphology and behavior. Proceedings of the National Academy of Sciences. 104 (2), 565-569.
3. Wang, M., Be´thou, O., Bradler, S., Jacques, FMB., Cui, Y., and Ren, D. (2014), Under Cover at Pre-Angiosperm Times: A Cloaked Phasmatodean Insect from the Early Cretaceous Jehol Biota. PLoS One, 9 (3), e91290
4. Farkas, TE., Mononen, T., Comeault, AA., Hanski, I. and Nosil, P. (2013) Evolution of Camouflage Drives Rapid Ecological Change in an Insect Community. Current Biology. 23, 1835-1843.
5. Nosil, P., Crespi, BJ., and Sandoval, CP. (2002) Host-plant adaptation drives the parallel evolution of reproductive isolation. Nature. 417, 440-443.
6. Nosil, P. and Crespi, BJ. (2006) Experimental evidence that predation promotes divergence in adaptive radiation. Proceedings of the National Academy of Sciences. 103 (24), 9090-9095.
7. Nosil, P. and Sandoval, CP. (2008) Ecological Niche Dimensionality and the Evolutionary Diversification of Stick Insects. PLoS One. 3(4), e1907
8. Whiting, MF., Bradler, S. and Maxwell, T. (2003) Loss and recovery of wings in stick insects. Nature. 421, 264-267.
9. Burrows, M. (2008) Jumping in a wingless stick insect, Timema chumash (Phasmatodea, Timematodea, Timematidae). The Journal of Experimental Biology. 211, 1021-1028.
10. Wolf, H., Bässler, U., Spieß, R. and Kittman, R. (2001) The femur–tibia control system in a proscopiid (Caelifera, Orthoptera): a test for assumptions on the functional basis and evolution of twig mimesis in stick insects. The Journal of Experimental Biology. 204, 3815-3822.
11. Scali, V. (2009) Stick insects: parthenogenesis, polyploidy and beyond. In: Life and Time: The Evolution of Life and its History. Cleup, Padova. 171-192
A few years ago I read an op-ed piece in the journal Nature that celebrated the potential demise of mosquitoes as scientists prepared to release genetically modified mosquitoes in Brazil in an attempt to eradicate populations carrying malaria. What most struck me about the piece was that the author concluded that mosquitoes performed no ecological function and that the world would be a better place without these pestiferous nuisances. This statement left me feeling a little uneasy. How certain could we actually be that mosquitoes performed absolutely no ecological function?
“Eradicating any organism would have serious consequences for ecosystems — wouldn’t it? Not when it comes to mosquitoes…”
In 2014 I listened to a podcast produced by Radiolab that reiterated the pointlessness of mosquitoes and again I wondered whether this could really hold entirely true. Apart from David Quammen’s valiant effort to convince us of the mosquito’s general innocence (it is after all only the females that bite, and even this is only in order to produce young). He also asks us to imagine just how quickly deforestation and exploitation of the tropics would have progressed without the relative protection afforded by the mosquito and all of it’s diseases.
According to the World Health Organisation (WHO) 17% of the global estimate of all infectious diseases are vector-borne. Of these, mosquito-borne diseases constitute the majority, with malaria causing an estimated 627,000 deaths in 2012 and infecting 1.5 to 2.7 million people a year. Some of the other mosquito-borne diseases that affect humans are Dengue fever, West Nile virus, Yellow fever, Lymphatic filariasis, Japanese encephalitis, Rift Valley fever, and Chikungunya; causing death, suffering and both social and economic hardship.
There are approximately 3,500 named mosquito species in the world. They are found in a variety of habitats in every biogeographic region apart from the Antarctic.Of these, only 40 Anopheles species are known to be effective transmitters of human malarial infection and only around 350 species are regarded as effective in all mosquito-borne human disease transmission. The catholic nature of mosquitoes in relation to habitat selectivity is best illustrated in the breadth of the geographic area covered by dominant malarial Anopheles mosquitoes. Mosquitoes are highly speciose, with the greatest species diversity being found in the Neotropical regions as shown in the map below.
This preponderance of mosquitoes to cause such human hardship has led to a variety of campaigns designed to control and eradicate them; from the use of DDT in the 1940s to attempts to sterilise males through exposure to radiation. Though there has been some success with these methods in the past, elimination of mosquitoes in the tropics has always proven difficult due to mosquito resistance, pathogen resistance to treatments, the lack of infrastructure and financial support. Conventional means of avoiding infection from mosquito-borne diseases have been to prevent being bitten through the use of mosquito nets and chemical repellents. I was therefore rather intrigued to hear about the work of Oxitec, the Alphey Lab and others in relation to developing genetic controls to exterminate this “winged scourge”.
The ecological niche filled by mosquitoes is little understood and has been poorly studied. In 2010, at the British Ecological Society’s annual meeting, the chair, Professor Charles Godfray said:
“We know very little about the [mosquito] community ecology… and this is significant because if you were to knock it out then you want to know what would take its place. […] And we don’t know enough, not for the want of trying, about the dispersal of the mosquitoes; how they move from one place to another.”
I simply couldn’t believe that such a large knowledge-gap existed with regards such an ubiquitous insect, so I decided to survey the scientific literature to figure out what is currently understood to be the ecological function performed by mosquitoes. I found that a very small number of papers actually concerned themselves with this topic directly and those that did were generally in relation to highly specific niches like larval processing of detritus chain interactions within pitcher-plants, the pollination of orchids, or focused on other species entirely, such as reindeer and caribou whose migration behaviour is influenced by the predation of mosquitoes and other biting flies. Understandably, most papers concentrated on the mosquito as disease vector – especially in relation to humans – but, apart from noting that mosquitoes constitute an enormous biomass, are found in both freshwater and terrestrial ecosystems at different life stages, and that they are highly speciose; there has been little scientific research into their ecological significance.We can extrapolate that they must be an important food source for a number of other insects, birds, reptiles, fish, amphibians and even mammals, but the data is lacking to support this – we need more research to be conducted to be certain. There is also a possibility that mosquitoes contribute to a disease dilution effect, but further study would be required to support any such claim.
So, is it a good idea to locally exterminate mosquitoes if we really don’t have any idea what will happen to their ecosystems? I would suggest that it probably isn’t the greatest idea. Possible scenarios are a reduction in available food for predators that will cause greater predation on other food sources thereby decreasing these at a faster rate and increasing competition. Increased competition can in turn lead to lower reproductive success and in the worst-case scenarios population collapse of apex predators. At least, I think it would be safe to assume that those ecosystems would no longer operate in the same way – their species composition would shift to a greater or lesser degree and with that the functional ecology.
And what about the disease dilution effect? Well, if it holds true in the case of mosquitoes then we may witness an intensification of disease virulence and higher infection rates. An alternative hypothesis is that the pathogens might move into other host species and we would be left scrabbling for new control mechanisms.
As someone with a desire to understand the intricately interlinked nature of our world and all the living creatures in it, I couldn’t support the deliberate extinction of any species (despite the detrimental effects it can have on humanity) without first knowing what the knock-on effects of that extinction would be. In doing something that we hope would benefit humanity, we may in fact be creating new and more complex problems.
An American entomologist, Jeremy Lockwood, wrote of the need to establish an ethical basis of “philosophically sound, scientifically consistent” considerations with regards our relationship to insects. He proposed that we refrain from taking actions that would kill or cause nontrivial pain to insects, but not if by avoiding those actions there would be nontrivial costs to human welfare. Genetically modified mosquitoes, and by association other mosquito control mechanisms, would presumably be considered acceptable to most people within this anthropocentric ethical framework. The irony of this position however is, as Lockwood points out, that a person considered a humanitarian is often referred to as, “one who wouldn’t hurt a fly”.
This blogpost is based on my final-year research paper. For those of you wanting a bit more in-depth information, you can read the full paper here.
References:
Alphey, L. (2014). Genetic Control of Mosquitoes. Annual Review of Entomology, 59(1), 205–224. doi:10.1146/annurev-ento-011613-162002
A selection of Sphingidae moths photographed from the collection at London’s Natural History Museum.
Commonly known as Hawk Moths, Sphinx Moths and Hornworms, these moths are important pollinators of orchids and other flowers.
Nectar tubes and hawk moth tongue lengths are often associated; Xanthopan morganii praedicta (centre) was famously predicted to exist by Charles Darwin and Alfred Russel Wallace based on the length of the nectar spur of the Madagascan Christmas star orchid (Angraecum sesquipedale). Note the length of the moth’s uncoiled proboscis in the photo above.
In a letter to Joseph Hooker in 1862, Darwin wrote:
“I have just received such a Box full […] with the astounding Angraecum sesquipedalia [sic] with a nectary a foot long. Good Heavens what insect can suck it.”
A hand-coloured lithograph of Angraceum sesquipedale by W.H. Fitch (1859) taken from Curtis’s Botanical Magazine.
A few days later, Darwin wrote a second letter in which he postulated that this insect must be a moth, and in 1867 Wallace published an article in which he supported Darwin’s suggestion, remarking that the African hawkmoth Xanthopan morganii (then known as Macrosila morganii) had a proboscis almost long enough to reach the bottom of the spur. In a footnote to this article Wallace wrote:
“That such a moth exists in Madagascar may be safely predicted; and naturalists who visit that island should search for it with as much confidence as astronomers searched for the planet Neptune,–and they will be equally successful!”
In 1903 (41 years after Darwin’s observation) this moth was discovered and named by Rothschild & Jordan after Wallace’s prediction that the moth would in fact be a hawkmoth. However, it was not before 1997 that it was finally confirmed that the Madagascan Christmas star orchid is actually pollinated by Xanthopan morganii praedicta.
Miller, W. E. (1997) ‘Diversity and evolution of tongue lengths in Hawkmoths (Sphingidae)’. Journal of the Lepidopterists’ Society. 51(1), 9-31
Wasserthal, L. T. (1997) ‘The pollinators of the Malagasy star orchids Angraecum sesquipedale, A. sororium and A. compactum and the evolution of extremely long spurs by pollinator shift’. Bot. Acta 110: 343-359